- Research Article
- Open access
- Published:
Global Existence and Convergence of Solutions to a Cross-Diffusion Cubic Predator-Prey System with Stage Structure for the Prey
Boundary Value Problems volume 2010, Article number: 285961 (2010)
Abstract
We study a cubic predator-prey system with stage structure for the prey. This system is a generalization of the two-species Lotka-Volterra predator-prey model. Firstly, we consider the asymptotical stability of equilibrium points to the system of ordinary differential equations type. Then, the global existence of solutions and the stability of equilibrium points to the system of weakly coupled reaction-diffusion type are discussed. Finally, the existence of nonnegative classical global solutions to the system of strongly coupled reaction-diffusion type is investigated when the space dimension is less than 6, and the global asymptotic stability of unique positive equilibrium point of the system is proved by constructing Lyapunov functions.
1. Introduction and Mathematical Model
The predator-prey model as, which follows, the ordinary differential equation system

is said to be the general Lotka-Volterra predator-prey model in [1–3], and to be cubic predator-prey system in [4], where  are the population densities of prey and predator species at time
are the population densities of prey and predator species at time  , respectively.
, respectively.  are positive constants,
are positive constants,  is nonnegative as the intrinsic growth rate of prey population, and the sign of
is nonnegative as the intrinsic growth rate of prey population, and the sign of  is undetermined.
is undetermined.  is the net mortality rate of predator population, and the survival of predator species is dependent on the survival state of prey species, and
is the net mortality rate of predator population, and the survival of predator species is dependent on the survival state of prey species, and  are the respective density restriction terms of prey and predator species.
are the respective density restriction terms of prey and predator species.  is the predation rate of the predator, and
is the predation rate of the predator, and  is the conversion rate of the predator. In [4], three questions about system (1.1) are discussed: the stability of nonnegative equilibrium points, and the existence, as well as numbers of limit cycle.
is the conversion rate of the predator. In [4], three questions about system (1.1) are discussed: the stability of nonnegative equilibrium points, and the existence, as well as numbers of limit cycle.
Referring to [5], we establish cubic predator-prey system with stage structure for the prey as follows:

where  and
and  are the population densities of the immature and mature prey species, respectively, and
are the population densities of the immature and mature prey species, respectively, and  denotes the density of the predator species. The predators live only on the immature prey species, as well as the survival of the predator species is dependent on the survival state of the immature prey species.
denotes the density of the predator species. The predators live only on the immature prey species, as well as the survival of the predator species is dependent on the survival state of the immature prey species.  are positive constants, and the sign of
are positive constants, and the sign of  is undetermined.
is undetermined.  and
and  are the birth rate and the mortality rate of the immature prey species, respectively.
are the birth rate and the mortality rate of the immature prey species, respectively.  and
and  are the net mortality rate of the mature prey population and the predator population, and
are the net mortality rate of the mature prey population and the predator population, and  is the conversion rate of the immature prey to the mature prey species.
is the conversion rate of the immature prey to the mature prey species.  and
and  are the respective density restriction terms of the immature prey species and predator species.
are the respective density restriction terms of the immature prey species and predator species.  is the predation rate of the predator to the immature prey population, and
is the predation rate of the predator to the immature prey population, and  is the conversion rate of the predator.
is the conversion rate of the predator.
Using the scaling

and redenoting  by
by  , system (1.2) reduces to
, system (1.2) reduces to

where  and
and  are positive constants, and
are positive constants, and  is undetermined to the sign.
is undetermined to the sign.
To take into account the inhomogeneous distribution of the predators and prey in different spatial locations within a fixed bounded domain  at any given time, and the natural tendency of each species to diffuse to areas of smaller population concentration, we derive the following PDE system of reaction-diffusion type:
at any given time, and the natural tendency of each species to diffuse to areas of smaller population concentration, we derive the following PDE system of reaction-diffusion type:

where  ,
,  is the unit outward normal vector of the boundary
is the unit outward normal vector of the boundary  which we will assume to be smooth. The homogeneous Neumann boundary condition indicates that the above system is self-contained with zero population flux across the boundary. The positive constants
which we will assume to be smooth. The homogeneous Neumann boundary condition indicates that the above system is self-contained with zero population flux across the boundary. The positive constants  ,
,  , and
, and  are said to be the diffusion coefficients, and the initial values
are said to be the diffusion coefficients, and the initial values  (
( ) are nonnegative smooth functions.
) are nonnegative smooth functions.
Note that, in recent years, there has been considerable interest to investigate the global behavior of a system of interacting populations by taking into account the effect of self as well as cross-diffusion. According to the ideas in [6–13], especially to [8, 9], the cross-diffusion term will be only included in the third equation, that is, the following cross-diffusion system:

In the above,  and
and  are positive constants.
are positive constants.  and
and  are the diffusion rates of the three species, respectively.
are the diffusion rates of the three species, respectively.  are referred to as self-diffusion pressures.
are referred to as self-diffusion pressures.  and
and  are cross-diffusion pressures. The term self-diffusion implies the movement of individuals from a higher to a lower concentration region. Cross-diffusion expresses the population fluxes of one species due to the presence of the other species. Generally, the value of the cross-diffusion coefficient may be positive, negative, or zero. The term positive cross-diffusion coefficient denotes the movement of the species in the direction of lower concentration of another species, and negative cross-diffusion coefficient denotes that one species tends to diffuse in the direction of higher concentration of another species [9].
are cross-diffusion pressures. The term self-diffusion implies the movement of individuals from a higher to a lower concentration region. Cross-diffusion expresses the population fluxes of one species due to the presence of the other species. Generally, the value of the cross-diffusion coefficient may be positive, negative, or zero. The term positive cross-diffusion coefficient denotes the movement of the species in the direction of lower concentration of another species, and negative cross-diffusion coefficient denotes that one species tends to diffuse in the direction of higher concentration of another species [9].
The main purpose of this paper is to study the asymptotic behavior of the solutions of the reaction-diffusion system (1.5) and the global existence of the solution of the cross-diffusion system (1.6). But it is necessary to denonstrate that the conclusion for the existence of global solution of system (1.6) in this paper is the generalization of the work to Lotka-Volterra competition model with cross-diffusion [11] and that the convergence of solution investigated in this paper which is not discussed in [11].
The paper will be organized as follows. In Section 2, we analyze the asymptotical stability of equilibrium points for the ODE system (1.4) via linearization and the Lyapunov method. In Section 3, we prove the global existence of solutions and the stability of the equilibrium points to the diffusion system (1.5). In Section 4, we investigate the existence of nonnegative classical global solutions by assuming  ,
,  ,
,  ,
,  ,
,  ,
,  to be positive constants only for the simplicity of calculation, and the global asymptotic stability of unique positive equilibrium point to the cross-diffusion system (1.6).
to be positive constants only for the simplicity of calculation, and the global asymptotic stability of unique positive equilibrium point to the cross-diffusion system (1.6).
2. Equilibrium Solution of the ODE System
In this section we discuss the stability of unique positive equilibrium point for system (1.4). The following theorem shows that the solution of system (1.4) is bounded.
Theorem 2.1.
Let  be the solution of system (1.4) with initial values
be the solution of system (1.4) with initial values  , and let
, and let  be the maximal existence interval of the solution. Then
be the maximal existence interval of the solution. Then  , where
, where

The above  is a positive constant depending only on
is a positive constant depending only on  , and further
, and further  .
.
Proof.
It is easy to see that (1.4) has a unique positive local solution  . Let
. Let  be the maximal existence time of the solution, and combin
be the maximal existence time of the solution, and combin  and
and  linearly, that is,
linearly, that is,  , it follows from (1.4) that
, it follows from (1.4) that

Using Young inequality, we can check that there exists a positive constant  depending only on
depending only on  and
and  such that
such that

It follows that

which implies that there exist  and
and  referring to (2.1) such that
referring to (2.1) such that  ,
,  , and
, and  .
.
Finally, we note that  . Let
. Let  , then
, then

From the comparison inequality for the ODE, we have  ,
,  .
.
Thus the solutions for system (1.4) are bounded. Further, from the extension theorem of solutions, we have  .
.
By the simple calculation, the sufficient conditions for system (1.4) having a unique positive equilibrium point as follows:
-
(i)
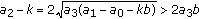 ; (ii)
; (ii) 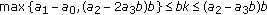 , where the left equal sign holds if and only if
, where the left equal sign holds if and only if 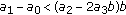 ; (iii)
; (iii) 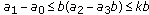 ; (iv)
; (iv)  ; (v)
; (v) 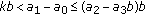 and
and 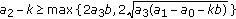 , where the second equal sign holds if and only if
, where the second equal sign holds if and only if 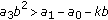 ; (vi)
; (vi) 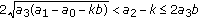 and
and 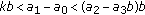 .
.
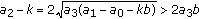 ; (ii)
; (ii) 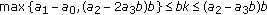 , where the left equal sign holds if and only if
, where the left equal sign holds if and only if 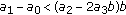 ; (iii)
; (iii) 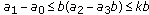 ; (iv)
; (iv)  ; (v)
; (v) 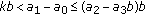 and
and 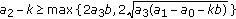 , where the second equal sign holds if and only if
, where the second equal sign holds if and only if 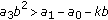 ; (vi)
; (vi) 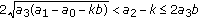 and
and 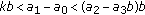 .
.If one of the above conditions holds, then system (1.4) has the unique positive equilibrium point  , where
, where

Theorem 2.2.
System (1.4) has the unique positive equilibrium point  when one of the above conditions (i), (ii), (iii), (iv), (v), and (vi) holds. If
when one of the above conditions (i), (ii), (iii), (iv), (v), and (vi) holds. If  holds, then
holds, then  is locally asymptotically stable.
is locally asymptotically stable.
Theorem 2.2 is easy to be obtained by using linearization; therefore, we omit its proof. The objective of this section is to prove the following result.
Theorem 2.3.
System (1.4) has the unique positive equilibrium point  when one of the above conditions (i), (ii), (iii), (iv), (v), and (vi) holds. If
when one of the above conditions (i), (ii), (iii), (iv), (v), and (vi) holds. If  holds, then
holds, then  is globally asymptotically stable.
is globally asymptotically stable.
Proof.
We make use of the general Lyapunov function

where  are positive constants. It holds that
are positive constants. It holds that  for any
for any  . Calculating the derivative along each solution of system (1.4), we have
. Calculating the derivative along each solution of system (1.4), we have

Let  and
and  . Then
. Then

We observe that

is a sufficient condition of  . So, when condition (2.10) holds, we have
. So, when condition (2.10) holds, we have

Set  . According to the Lyapunov-LaSalle invariance principle [14],
. According to the Lyapunov-LaSalle invariance principle [14],  is global asymptotic stability if inequality (2.10) and all conditions of Theorem 2.2 are satisfied. Theorem 2.3 is, thus, proved.
is global asymptotic stability if inequality (2.10) and all conditions of Theorem 2.2 are satisfied. Theorem 2.3 is, thus, proved.
3. Stability of the PDE System without Cross-Diffusion
In this section, we first prove the global existence and uniform boundedness of solutions, then discuss the stability of unique positive equilibrium solution for the weakly coupled reaction-diffusion system (1.5).
Denote that  , where
, where  and
and  . It is easy to see that
. It is easy to see that  with
with  . The standard PDE theory [15] shows that (1.5) has the unique solution
. The standard PDE theory [15] shows that (1.5) has the unique solution  , where
, where  is the maximal existence time. The following theorem shows that the solution of (1.5) is uniformly bounded, and thus
is the maximal existence time. The following theorem shows that the solution of (1.5) is uniformly bounded, and thus  .
.
Theorem 3.1.
Let  be the solution of system (1.5) with initial values
be the solution of system (1.5) with initial values  , and let
, and let  be the maximal existence time. Then
be the maximal existence time. Then  ,
,  , and
, and  , where
, where  is a positive constant depending only on
is a positive constant depending only on  and all coefficients of (1.5) and
and all coefficients of (1.5) and  ,
,  . Furthermore,
. Furthermore,  and
and  on
on  for any
for any  if
if  .
.
Proof.
Let  be the solution of (1.5) with initial values
be the solution of (1.5) with initial values  . From the maximum principle for parabolic equations [16], it is not hard to verify that
. From the maximum principle for parabolic equations [16], it is not hard to verify that  for
for  , where
, where  is the maximal existence time of the solution
is the maximal existence time of the solution  . Furthermore, we know by the strong maximum principle that
. Furthermore, we know by the strong maximum principle that  on
on  for all
for all  if
if  . Next we prove that the solution
. Next we prove that the solution  is bounded on
is bounded on  .
.
Integrating the first two equations of (1.5) over  and adding the results linearly, we have that, by Young inequality,
and adding the results linearly, we have that, by Young inequality,

for some positive constant  depending only on the coefficients of (1.5). Therefore,
depending only on the coefficients of (1.5). Therefore,  is bounded in
is bounded in  . Using [17, Exercise 5 of Section
. Using [17, Exercise 5 of Section  ], we obtain that
], we obtain that  is also bounded in
is also bounded in  . Now note that
. Now note that  The maximum principle gives
The maximum principle gives  . The proof of Theorem 3.1 is completed.
. The proof of Theorem 3.1 is completed.
In order to prove the global stability of unique positive equilibrium solution for system (1.5), we first recall the following lemma which can be found in [7, 17].
Lemma 3.2.
Let  and
and  be positive constants. Assume that
be positive constants. Assume that  ,
,  ,
,  and
and  is bounded from below. If
is bounded from below. If  , and
, and  in
in  for some constant
for some constant  , then
, then 
Let  be the eigenvalues of the operator
be the eigenvalues of the operator  on
on  with the homogeneous Neumann boundary condition, and let
with the homogeneous Neumann boundary condition, and let  be the eigenspace corresponding to
be the eigenspace corresponding to  in
in  . Denote that
. Denote that  ,
,  is an orthonormal basis of
is an orthonormal basis of  and
and  . Then
. Then

Next we present the clear proof of the the global stability by two steps:
Step 1 (Local Stability).
Let  and
and  , where
, where

The linearization of (1.5) at  is
is

For each  ,
,  is invariant under the operator
is invariant under the operator  , and
, and  is an eigenvalue of
is an eigenvalue of  on
on  if and only if it is an eigenvalue of the matrix
if and only if it is an eigenvalue of the matrix  .
.
The characteristic polynomial of  is given by
is given by

where

Thus

where  and
and  are given by
are given by

According to the Routh-Hurwitz criterion [18], for each  , the three roots
, the three roots  of
of  all have negative real parts if and only if
all have negative real parts if and only if  ,
,  and
and  . Noting that
. Noting that  and
and  , the three roots have negative real parts if
, the three roots have negative real parts if  . A direct calculation shows that
. A direct calculation shows that  is negative if
is negative if

Now we can conclude that there exists a positive constant  such that
such that

In fact, let  , then
, then

Since  as
as  , it follows that
, it follows that

It is easy to see that  are the three roots of
are the three roots of  . Thus, there exists a positive constant
. Thus, there exists a positive constant  such that
such that

By continuity, we see that there exists  such that the three roots
such that the three roots  of
of  satisfy
satisfy

So

Let

then  , and (3.10) holds for
, and (3.10) holds for  .
.
Consequently, the spectrum of  , consisting only of eigenvalues, lies in
, consisting only of eigenvalues, lies in  if (3.9) holds, and the local stability of
if (3.9) holds, and the local stability of  follows [19, Theorem
follows [19, Theorem  ].
].
Step 2 (Global Stability).
In the following,  denotes a generic positive constant which does not depend on
denotes a generic positive constant which does not depend on  and
and  . Let
. Let  be the unique positive solution. Then it follows from Theorem 3.1 that
be the unique positive solution. Then it follows from Theorem 3.1 that  is bounded uniformly on
is bounded uniformly on  , that is,
, that is,  for all
for all  . By [20, Theorem
. By [20, Theorem  ],
],

Define the Lyapunov function

Then  for all
for all  . Using (1.5) and integrating by parts, we have
. Using (1.5) and integrating by parts, we have

Taking  and
and  , we have that
, we have that

where  holds for
holds for

From Theorem 3.1 the solution  of (1.5) is bounded, and so are the derivatives of
of (1.5) is bounded, and so are the derivatives of  and
and  by equations in (1.5). Applying Lemma 3.2, we obtain
by equations in (1.5). Applying Lemma 3.2, we obtain

As  , it follows that
, it follows that

Using inequality (3.17) and system (1.5), the derivative of  is bounded in
is bounded in  . From Lemma 3.2, we conclude that
. From Lemma 3.2, we conclude that  as
as  . Therefore
. Therefore

Using the Poincaré inequality yields

where  Thus, it follows from (3.22) and (3.25) that
Thus, it follows from (3.22) and (3.25) that

as  . So we have
. So we have  as
as  . Similarly,
. Similarly,  as
as  . Therefore, there exists a sequence
. Therefore, there exists a sequence  with
with  such that
such that  . As
. As  is bounded, there exists a subsequence of
is bounded, there exists a subsequence of  , still denoted by the same notation, and nonnegative constant
, still denoted by the same notation, and nonnegative constant  such that
such that

At  , from the first equation of (1.5), we have
, from the first equation of (1.5), we have

In view of (3.22) and (3.27), it follows from (3.28) that  , thus
, thus

According to (3.17), there exists a subsequence of  , denoted still by
, denoted still by  , and nonnegative functions
, and nonnegative functions  , such that
, such that

In view of (3.29) and noting that in fact  and
and  , we know that
, we know that  . Therefore,
. Therefore,

The global asymptotic stability of  follows from (3.31) and the local stability of
follows from (3.31) and the local stability of  .
.
Theorem 3.3.
System (1.5) has the unique positive equilibrium point  when one of the conditions (i), (ii), (iii), (iv), (v), and (vi) in Section 2 holds. If (3.9) and (3.21) hold, then
when one of the conditions (i), (ii), (iii), (iv), (v), and (vi) in Section 2 holds. If (3.9) and (3.21) hold, then  is globally asymptotically stable.
is globally asymptotically stable.
4. Global Existence of Classical Solutions and Convergence
In this section, we discuss the existence of nonnegative classical global solutions and the global asymptotic stability of unique positive equilibrium point of system (1.6).
Some notations throughout this section are as follows:  ,
,  means that
means that  for any
for any  with
with  ,
,  ,
,  means that
means that  and
and  are in
are in  ,
,  and
and  with
with  .
.
To obtain  normal estimates of the solution for (1.6), we present a series of lemmas in the following.
normal estimates of the solution for (1.6), we present a series of lemmas in the following.
Lemma 4.1.
Let  be the solution of (1.6). Then there exists a positive constant
be the solution of (1.6). Then there exists a positive constant  (
( 1) such that
1) such that

Proof.
By applying the comparison principle [20] to system (1.6), we have  and
and  in
in  . To prove that
. To prove that  in the following, we consider the auxiliary problem
in the following, we consider the auxiliary problem

Notice that the functions  and
and  are sufficiently smooth in
are sufficiently smooth in  , and are quasimonotone in
, and are quasimonotone in  . Let
. Let  and
and  be a pair of upper-lower solutions for (4.2), where
be a pair of upper-lower solutions for (4.2), where  and
and  are positive constants. Direct calculation with inequalities
are positive constants. Direct calculation with inequalities

yields  and
and  . It follows that there exists
. It follows that there exists  for any
for any  , where
, where  is a big enough positive constant such that (4.1) holds.
is a big enough positive constant such that (4.1) holds.
Lemma 4.2.
Let  , and
, and  for the solution to following equation:
for the solution to following equation:

where  ,
,  are positive constants and
are positive constants and  . Then there exists a positive constant
. Then there exists a positive constant  , depending on
, depending on  and
and  , such that
, such that

Furthermore,

Proof.
It is easy to check, from  , that
, that

where  and
and  .
.  and
and  are bounded in
are bounded in  from (4.1). Multiplying (4.7) by
from (4.1). Multiplying (4.7) by  , and integrating by parts over
, and integrating by parts over  , yields
, yields

Using Hölder inequality and Young inequality to estimate the right side of (4.8), we have

with some  . Substituting (4.9) into (4.8) yields
. Substituting (4.9) into (4.8) yields

where  depends on
depends on  and
and  . Since
. Since  , the elliptic regularity estimate [10, Lemma
, the elliptic regularity estimate [10, Lemma  ] yields
] yields

From (4.7), we have  . Hence,
. Hence,  . Moreover, the Sobolev embedding theorem shows that (4.6) holds.
. Moreover, the Sobolev embedding theorem shows that (4.6) holds.
Lemma 4.3 (Lemma  can be presented by combining Lemmas
can be presented by combining Lemmas  and
and  in [11]).
in [11]).
Let  , and let
, and let  satisfy
satisfy

and there exist positive constants  and
and  such that
such that  . Then there exists a positive constant
. Then there exists a positive constant  independent of
independent of  but possibly depending on
but possibly depending on  ,
,  ,
,  ,
,  and
and  such that
such that

Finally, one proposes some standard embedding results which are important to obtain the  normal estimates of the solution for (1.6).
normal estimates of the solution for (1.6).
Lemma 4.4.
Let  be a fixed bounded domain and
be a fixed bounded domain and  . Then for all
. Then for all  with
with  , one has
, one has
(1) 
(2) 
(3) 
where  is a positive constant dependent on
is a positive constant dependent on  and
and  .
.
The main result about the global existence of nonnegative classical solution for the cross-diffusion system (1.6) is given as follows.
Theorem 4.5.
Assume that  and
and  satisfy homogeneous Neumann boundary conditions and belong to
satisfy homogeneous Neumann boundary conditions and belong to  for some
for some  . Then system (1.6) has a unique nonnegative solution
. Then system (1.6) has a unique nonnegative solution  when the space dimension is
when the space dimension is  .
.
Proof.
Step 1.
 -
- -Estimates and
-Estimates and -Estimates of
-Estimates of . Firstly, integrating the third equation of (1.6) over
. Firstly, integrating the third equation of (1.6) over  , we have
, we have

Thus

Furthermore

Integrating (4.14) in  and moving terms yield
and moving terms yield

Secondly, multiplying the third equation of (1.6) by  and integrating over
and integrating over  , we have
, we have

Integrating the above expression in  yields
yields

Since  from Lemma 4.2, and using Hölder inequality and Young inequality, we have
from Lemma 4.2, and using Hölder inequality and Young inequality, we have

From (4.1) and  , it holds that
, it holds that

Taking  and selecting a proper
and selecting a proper  such that
such that  , then applying (4.20) and (4.21) to (4.19) yields
, then applying (4.20) and (4.21) to (4.19) yields

Denote that  . Then it follows from (4.22) that
. Then it follows from (4.22) that

It is easy to see that  and
and  for any
for any  ; hence
; hence

Take  . Then it follows from
. Then it follows from  -estimates of
-estimates of  namely (4.15), that
namely (4.15), that

It follows from Lemma 4.3 and (4.24) that

Since  ,
,  is bounded by contrary proof. It follows that
is bounded by contrary proof. It follows that  is bounded, that is,
is bounded, that is,  . It is easy to check that
. It is easy to check that  for all
for all  still denote
still denote  by
by  , then
, then

Finally, we observe that  satisfies
satisfies  with
with  . So take
. So take  for (4.17) and (4.19). Then there exists a positive constant
for (4.17) and (4.19). Then there exists a positive constant  such that
such that

Step 2.
 -Estimates of
-Estimates of . We rewrite the third equation of (1.6) as a linear parabolic equation
. We rewrite the third equation of (1.6) as a linear parabolic equation

where  ,
,  ,
,  are Kronecker symbols.
are Kronecker symbols.
To apply the maximum principle [15, Theorem  , page 181] to (4.15) to obtain
, page 181] to (4.15) to obtain  , we need to verify that the following conditions hold: (1)
, we need to verify that the following conditions hold: (1)  is bounded; (2)
is bounded; (2)  (3)
(3)  , where
, where  and
and  are positive constants, and
are positive constants, and  and
and  satisfy
satisfy

Next we verify conditions (1)–(3) in turn. From (4.28), condition (1) is true for  . One can choose
. One can choose  such that condition (2) holds. To verify condition (3), the first equation of (1.6) is written in the divergence form
such that condition (2) holds. To verify condition (3), the first equation of (1.6) is written in the divergence form

where  is bounded in
is bounded in  by Lemma 4.1, and
by Lemma 4.1, and  for
for  from (4.27). Application of the Hölder continuity result [15, Theorem
from (4.27). Application of the Hölder continuity result [15, Theorem  , page 204] to (4.19) yields
, page 204] to (4.19) yields

Returning to (4.7), since  for any
for any  by (4.1) and (4.27), and
by (4.1) and (4.27), and  by (4.32), then by applying the parabolic regularity theorem [15, Theorem
by (4.32), then by applying the parabolic regularity theorem [15, Theorem  , pages 341-342] to (4.7) we have
, pages 341-342] to (4.7) we have

Hence  from Lemma 4.4, which shows that
from Lemma 4.4, which shows that  . Similarly,
. Similarly,  by the second equation of (1.6). Now we can show that
by the second equation of (1.6). Now we can show that  , which imply that
, which imply that  . In addition,
. In addition,  obviously belongs to
obviously belongs to  . It follows that one can select
. It follows that one can select  . Now the above three conditions are satisfied, and
. Now the above three conditions are satisfied, and  from [15, Theorem
from [15, Theorem  , page 181]. Recalling Lemma 4.1, thus there exists a positive constant
, page 181]. Recalling Lemma 4.1, thus there exists a positive constant  for any
for any  such that
such that

Step 3.
The Proof of the Classical Solution of (1.6) in
of (1.6) in for Any
for Any . Because
. Because  , we have from (4.34) that
, we have from (4.34) that  for any
for any  . So
. So  for all
for all  . It follows from [15, Lemma
. It follows from [15, Lemma  , page 80] that
, page 80] that  . And direct calculation
. And direct calculation  yields
yields  . So we have
. So we have

The third equation of (1.6) can be written as

Summarizing the above conclusions that are proved, we know that  and
and  are all bounded in
are all bounded in  . It follows from [15, Theorem
. It follows from [15, Theorem  page 204] that there exists
page 204] that there exists  such that
such that

The proof of Lemma 4.2 is similar. Then we have  , that is,
, that is,  . Applying the [13, Theorem
. Applying the [13, Theorem  page 204] to the second equation (1.6), there exists
page 204] to the second equation (1.6), there exists  such that
such that

Furthermore, applying Schauder estimate [15, page 320-321] yields  for
for  . Selecting
. Selecting  and using Sobolev embedding theorem, we have
and using Sobolev embedding theorem, we have  . Still applying Schauder estimate, we have
. Still applying Schauder estimate, we have

Let  . Then
. Then  satisfies
satisfies

where  . By (4.35)–(4.38), we have
. By (4.35)–(4.38), we have  . So applying Schauder estimate to (4.40) yields
. So applying Schauder estimate to (4.40) yields  . Since
. Since  , we have
, we have

The first equation of (1.6) can be written as

where  . By (4.35), (4.39), and (4.41), we have
. By (4.35), (4.39), and (4.41), we have  . So applying Schauder estimate to (4.42) yields
. So applying Schauder estimate to (4.42) yields

In particular, if  , then
, then  ; in other words, Theorem 4.5 is proved. For the case
; in other words, Theorem 4.5 is proved. For the case  , from Sobolev embedding theorem, we have
, from Sobolev embedding theorem, we have  . Repeating the above bootstrap and Shauder estimate arguments, this completes the proof of Theorem 4.5. About space dimension
. Repeating the above bootstrap and Shauder estimate arguments, this completes the proof of Theorem 4.5. About space dimension  , see [21].
, see [21].
Theorem 4.6.
System (1.6) has the unique positive equilibrium point  when one of the conditions (i), (ii), (iii), (iv), (v), and (vi) in Section 2 holds. Let the space dimension be
when one of the conditions (i), (ii), (iii), (iv), (v), and (vi) in Section 2 holds. Let the space dimension be  , and let the initial values
, and let the initial values  be nonnegative smooth functions and satisfy the homogenous Neumann boundary conditions. If the following condition (4.44) holds, then the solution
be nonnegative smooth functions and satisfy the homogenous Neumann boundary conditions. If the following condition (4.44) holds, then the solution  of (1.6) converges to
of (1.6) converges to  in
in  :
:

where  and
and  .
.
Proof.
Define the Lyapunov function

where  and
and  have been given in Theorem 4.6. Obviously,
have been given in Theorem 4.6. Obviously,  is nonnegative, and
is nonnegative, and  if and only if
if and only if  and
and  . When
. When  is a positive solution of system (1.6),
is a positive solution of system (1.6),  is well posed for all
is well posed for all  from Theorem 4.5. According to system (1.6), the time derivative of
from Theorem 4.5. According to system (1.6), the time derivative of  satisfies
satisfies

It is easy to check that the final three integrands on the right side of the above expression are positive definite because of the electing of  , and the sufficient and necessary conditions of the first integrand being positive definite are the following:
, and the sufficient and necessary conditions of the first integrand being positive definite are the following:

Noticing that (4.44) is the sufficient conditions of (4.47), so there exists a positive constant  such that
such that

Similar to the tedious calculations of  , using integration by parts, Hölder inequality, and (4.34), one can verify that
, using integration by parts, Hölder inequality, and (4.34), one can verify that  is bounded from above. Thus we have from (4.48) and Lemma 3.2 in Section 3 that
is bounded from above. Thus we have from (4.48) and Lemma 3.2 in Section 3 that

In addition,  is decreasing for
is decreasing for  , so we can conclude that the solution
, so we can conclude that the solution  is globally asymptotically stable. The proof of Theorem 4.6 is completed.
is globally asymptotically stable. The proof of Theorem 4.6 is completed.
References
Kuno E: Mathematical models for predator-prey interaction. Advances in Ecological Research 1987, 16: 252-265.
Zheng JB, Yu ZX, Sun JT: Existence and uniqueness of limit cycle for prey-predator systems with sparse effect. Journal of Biomathematics 2001,16(2):156-161.
Shen C, Shen BQ: A necessary and sufficient condition of the existence and uniqueness of the limit cycle for a class of prey-predator systems with sparse effect. Journal of Biomathematics 2003,18(2):207-210.
Huang X, Wang Y, Zhu L: One and three limit cycles in a cubic predator-prey system. Mathematical Methods in the Applied Sciences 2007,30(5):501-511. 10.1002/mma.791
Zhang X, Chen L, Neumann AU: The stage-structured predator-prey model and optimal harvesting policy. Mathematical Biosciences 2000,168(2):201-210. 10.1016/S0025-5564(00)00033-X
Shigesada N, Kawasaki K, Teramoto E: Spatial segregation of interacting species. Journal of Theoretical Biology 1979,79(1):83-99. 10.1016/0022-5193(79)90258-3
Pang PYH, Wang M: Strategy and stationary pattern in a three-species predator-prey model. Journal of Differential Equations 2004,200(2):245-273. 10.1016/j.jde.2004.01.004
Kuto K: Stability of steady-state solutions to a prey-predator system with cross-diffusion. Journal of Differential Equations 2004,197(2):293-314. 10.1016/j.jde.2003.10.016
Dubey B, Das B, Hussain J: A predator-prey interaction model with self and cross-diffusion. Ecological Modelling 2001,141(1–3):67-76.
Lou Y, Ni W-M, Wu Y: On the global existence of a cross-diffusion system. Discrete and Continuous Dynamical Systems 1998,4(2):193-203.
Choi YS, Lui R, Yamada Y: Existence of global solutions for the Shigesada-Kawasaki-Teramoto model with strongly coupled cross-diffusion. Discrete and Continuous Dynamical Systems 2004,10(3):719-730.
Zhang R, Guo L, Fu SM:Global behavior for a diffusive predator-prey model with stage-structure and nonlinear density restriction-II: the case in
 . Boundary Value Problems 2009, 2009:-19.
. Boundary Value Problems 2009, 2009:-19.Zhang R, Guo L, Fu SM:Global behavior for a diffusive predator-prey model with stage-structure and nonlinear density restriction-I: the case in
 . Boundary Value Problems 2009, 2009:-26.
. Boundary Value Problems 2009, 2009:-26.Hale JK: Ordinary Differential Equations. 2nd edition. Robert E. Krieger, Malabar, Fla, USA; 1980:xvi+361.
Ladyzenskaja OA, Solonnikov VA, Uralceva NN: Linear and Quasilinear Partial Differential Equations of Parabolic Type, Translations of Mathematical Monographs. Volume 23. American Mathematical Society, Providence, RI, USA; 1968.
Protter MH, Weinberger HF: Maximum Principles in Differential Equations. 2nd edition. Springer, New York, NY, USA; 1984.
Wang MX: Nonlinear Partial Differential Equations of Parabolic Type. Science Press, Beijing, China; 1993.
May RM: Stability and Complexity in Model Ecosystems. Princeton Univesity Press, Princeton, NJ, USA; 1974.
Henry D: Geometric Theory of Semilinear Parabolic Equations, Lecture Notes in Mathematics. Volume 840. Springer, Berlin, Germany; 1993.
Protter MH, Weinberger HF: Maximum Principles in Differential Equations. 2nd edition. Springer, New York, NY, USA; 1984:x+261.
Cao HH, Fu SM: Global solutions for a cubic predator-prey cross-diffusion system with stage structure. Mathematics in Practice and Theory 2008,38(21):161-177.
 . Boundary Value Problems 2009, 2009:-19.
. Boundary Value Problems 2009, 2009:-19. . Boundary Value Problems 2009, 2009:-26.
. Boundary Value Problems 2009, 2009:-26.Acknowledgments
The work of this author was partially supported by the Natural Science Foundation of Anhui Province Education Department (KJ2009B101) and the NSF of Chizhou College (XK0833) (caguhh@yahoo.com.cn). The work of this author was partially supported by the China National Natural Science Foundation (10871160), the NSF of Gansu Province (096RJZA118), and NWNU-KJCXGC-03-47, 61 Foundations (fusm@nwnu.edu.cn).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Cao, H., Fu, S. Global Existence and Convergence of Solutions to a Cross-Diffusion Cubic Predator-Prey System with Stage Structure for the Prey. Bound Value Probl 2010, 285961 (2010). https://doi.org/10.1155/2010/285961
Received:
Accepted:
Published:
DOI: https://doi.org/10.1155/2010/285961